












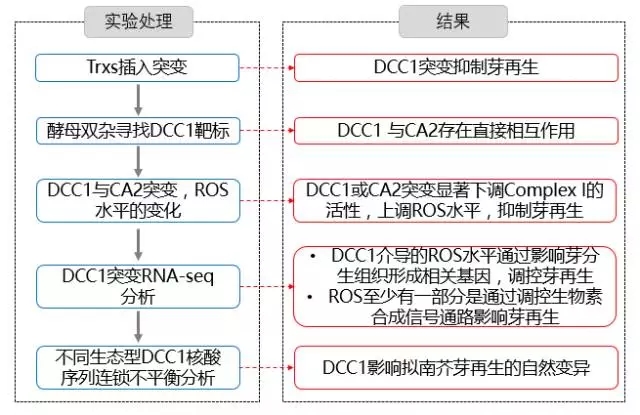
基迪奥合作客户文章-------植株再生的调控机制 返回
今年7月,山东农业大学张宪省教授在《Plant Physiology》杂志发表了一篇题为“Thioredoxin-mediated ROS Homeostasis Explains Natural Variation in Plant Regeneration”的文章,阐明了影响拟南芥再生能力自然变异的分子机制,基迪奥有幸参与该项目,配合完成了拟南芥野生型和突变体的转录组测序及生物信息学分析,筛选了影响芽再生的关键基因与信号通路。

合作单位:山东农业大学张宪省教授 发表杂志:《Plant Physiology》 影响因子:6.456
研究背景
植物再生对基因研究和农业应用是必需的,植物的再生能力在不同基因型中变化很大,但是造成这种变化的机制目前还没有被阐明。活性氧(ROS)是关键的信号分子,可以通过氧化翻译后修饰改变靶标蛋白活性,这些靶标蛋白参与激素信号感知与响应、MAPK信号转导等。
在根生长过程中,ROS内稳态对于细胞增殖到细胞分化的转变是至关重要的,而ROS内稳态被多种抗氧化因子调控,硫氧还蛋白(Trxs)就是清除ROS的关键因子。线粒体呼吸复合物I(the mitochondrial respiratory complex I,Complex I)亚基的氧化还原状态影响复合物的活性,碳酸酐酶2(CARBONIC ANHYDRASE 2, CA2)和另四个碳酸酐酶组成了一个Complex I功能性亚基,而Complex I的很多亚基是Trxs的潜在靶标。
实验材料与方法
对野生型拟南芥(Col-0-A, Col-0-B, Col-0-C)和硫氧还蛋白DCC1突变型拟南芥dcc1 (dcc1-A, dcc1-B, dcc1-C)进行RNA-seq分析,寻找芽再生关键基因。
对48个不同生态型拟南芥中的DCC1核酸序列进行连锁不平衡分析,寻找与芽再生相关的SNP位点。
研究思路
研究结果
1、DCC1突变抑制拟南芥芽再生能力
为了确定Trxs 在芽再生中的功能,利用T-DNA 插入突变,发现七种Trxs中只有AT5G50100 显著影响了芽再生,命名为DDC1。DCC1突变下调拟南芥的芽再生能力,包括低的芽分化频率和愈伤组织芽数量,而恢复突变后可以恢复表型(图1)。原位杂交和beta-GUS分析结果显示,在芽再生过程中,DCC1主要在愈伤组织内部区域表达。
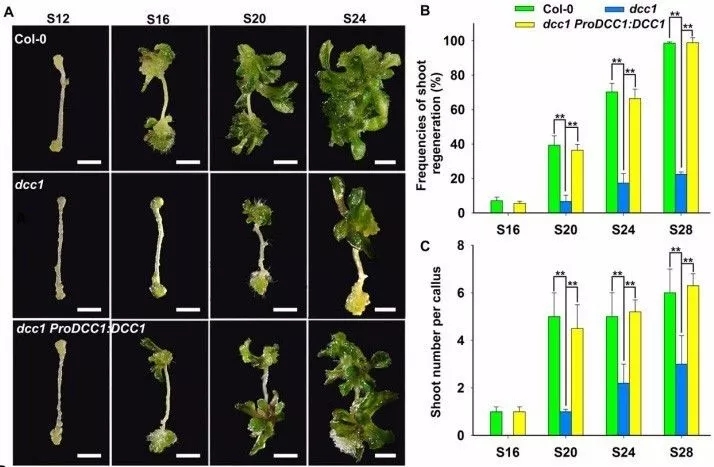
图1 DCC1突变影响芽再生能力
2、DCC1 编码功能性硫氧还蛋白
DCC1在功能未知的DUF393结构域的N端包含一个DxxCxxC基序(图2A),DxxCxxC基序在不同物种的DCC家族蛋白中具有序列保守性(图2B),进化分析结果显示DCC1和其他物种的蛋白均有高度同源性,如亚麻芥(Camelina sativa)、芜菁(Brassica rapa)和萝卜(Raphanus sativus)(图2C)。
两个半胱氨酸残基暗示DCC1调控二硫键交换,FiTC-insulin对Trx活性高度敏感,二硫键减少后显示更高的荧光表达,分析结果显示DCC1蛋白孵育后具有很高的荧光密度(图2D),表明DCC1作为Trx行使功能,并且发现其定位在线粒体(图2E)。
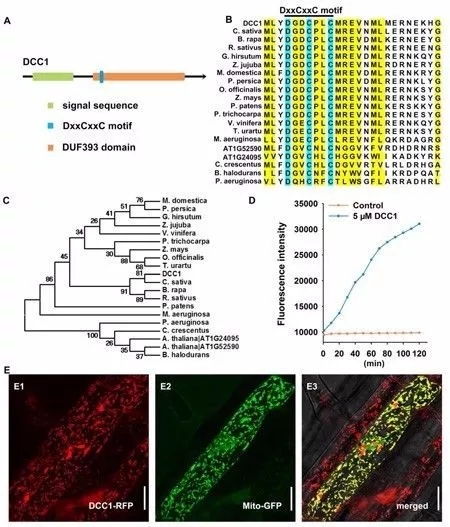
图2 DCC1 编码功能性硫氧还蛋白
3、DCC1或CA2突变显著下调Complex I的活性,上调ROS水平
酵母双杂试验、pull down、荧光素酶互补图像检测和DCC1-GFP免疫共沉淀试验表明DCC1和CA2存在直接相互作用(图3)。CA2突变显著下调芽再生能力,包括低的芽分化频率和愈伤组织的芽数量,而恢复突变后可以恢复表型。
DCC1或CA2突变显著下调Complex I的活性,同时突变DCC1、 CA2与单个突变没有更明显变化(图4A),表明这两种蛋白在相同的通路上起作用,且双突变的complex I活性可以被同时过表达DCC1和CA2蛋白完全回复(图4B)。利用DAB染色分析检测愈伤组织ROS水平发现,在芽再生过程中,DCC1或CA2突变上调愈伤组织ROS水平(图4C)。
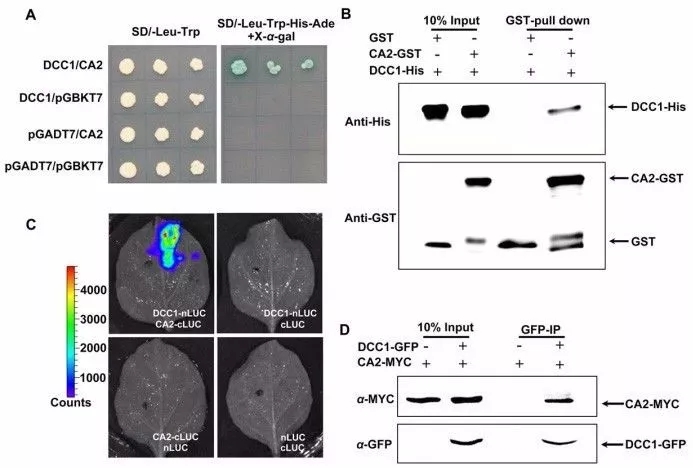
图3 DCC1与CA2直接相互作用
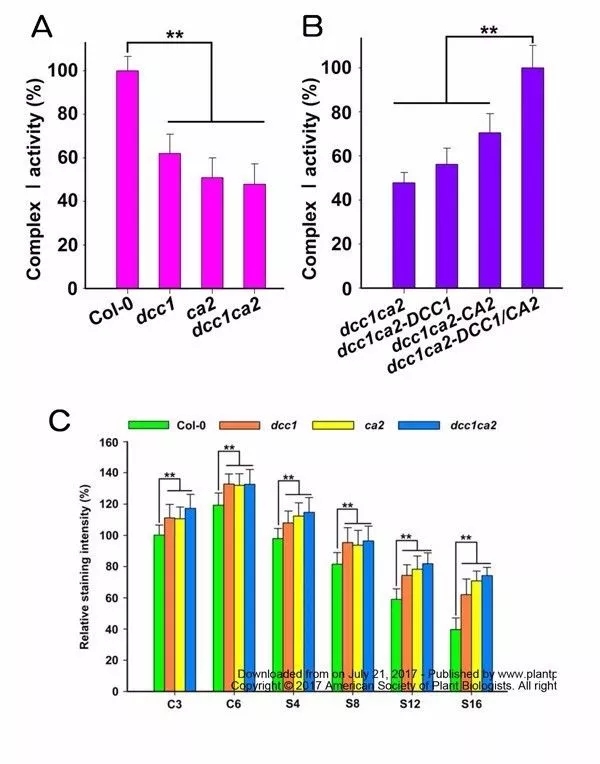
图4 DCC1或CA2突变影响线粒体呼吸复合物I的活性和ROS水平
4、ROS水平影响芽再生能力
H2O2能上调ROS水平,利用H2O2处理野生型外植体发现,H2O2抑制芽再生能力,并且浓度越高,抑制效果越显著(图5)。谷胱甘肽(GSH)是主要的内源清除ROS的抗氧化因子,DCC1、CA2突变后,GSH处理组相比未处理组,芽再生能力显著上调(图6)。过氧化氢酶3(CATALASE 3,CAT3)编码蛋白催化H2O2水解生成H2O和O2,过表达CAT3上调芽再生能力,过表达CAT3引起的ROS下调可以恢复突变的表型(图6),表明ROS水平对调节芽再生能力是至关重要的。
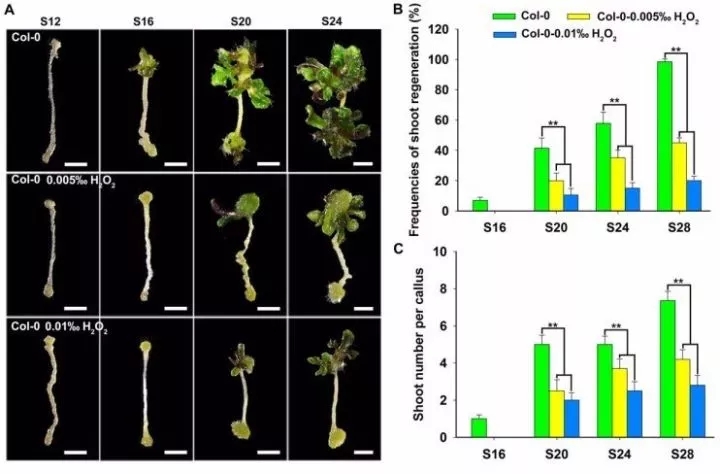
图5 ROS水平影响芽再生能力
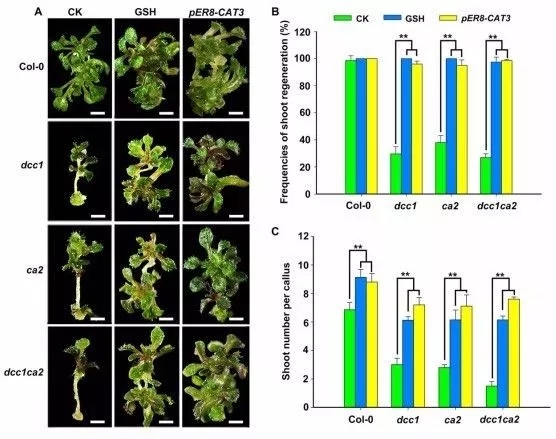
图6 GSH和CAT3对芽再生的影响
5、ROS 通过调控多种信号通路影响芽再生
野生型拟南芥和DCC1突变型拟南芥RNA-seq分析结果发现,在突变组中,689个基因显著上调,2332个基因显著下调(图7A)。GO富集分析显示DCC1调控多种生物学过程,比如生物素合成过程、芽系统发生、氧化还原过程、细胞分化及器官形态发生等(图7B)。
除了WUS和STM,分生组织主效基因KNAT1、KNAT2、KNAT4、WOXs、CUCs在DCC1突变体中也显著下调(图7C)。qRT-PCR结果验证了这些基因的转录水平,表明DCC1介导的ROS水平通过影响芽分生组织形成相关基因调控芽再生。
七个生物素合成基因和生物素响应基因在DCC1突变体中显著下调,利用已知的生物素标记分子DR5rev:GFP检测在芽再生过程中生物素响应信号分配,发现H2O2处理后GFP信号减弱,ROS抑制生物素信号,表明ROS至少有一部分是通过调控生物素合成信号通路影响芽再生。
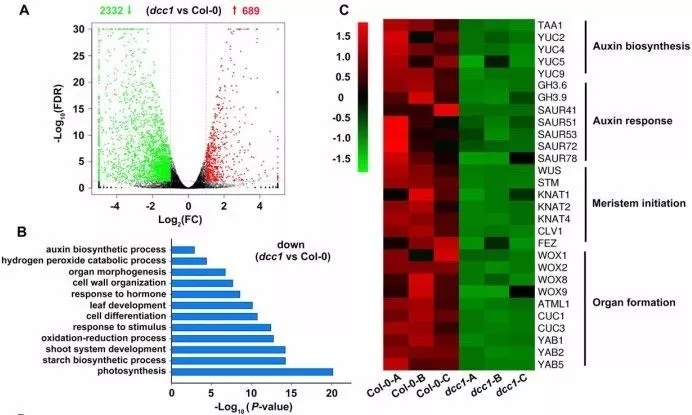
图7 野生型拟南芥和DCC1突变型拟南芥RNA-seq分析
6、DCC1 在拟南芥芽再生的自然变异中扮演重要角色
利用48个不同生态型中的DCC1核酸序列进行连锁不平衡分析,发现了六个SNPs (SNP 108、147、174、 175、378和 481)和芽再生频率高度相关,两个单倍型(Hap1和Hap2)确定了48个拟南芥的自然变异,Hap1包含30个生态型,Hap2包含18个生态型(图8A、B、C),Hap1生态型的芽再生频率和单个愈伤组织的芽数量高于Hap2生态型(图8D、E)。
六个SNPs中只有两个(SNP175和481)造成有义突变,175和481位点的突变没有影响DCC1的硫氧还蛋白活性(图8F、G),但是两个位点突变时DCC1和CA2没有相互作用,无法回复芽再生能力(图8H、I),表明DCC1的SNP175和SNP481对芽再生是关键的。
利用DAB染色检测Hap1和Hap2生态型的ROS水平发现,Hap2生态型的ROS水平显著高于Hap1,但芽再生能力低于Hap1(图9),表明DCC1介导的ROS体内稳态平衡对于拟南芥芽再生的自然变异是至关重要的。
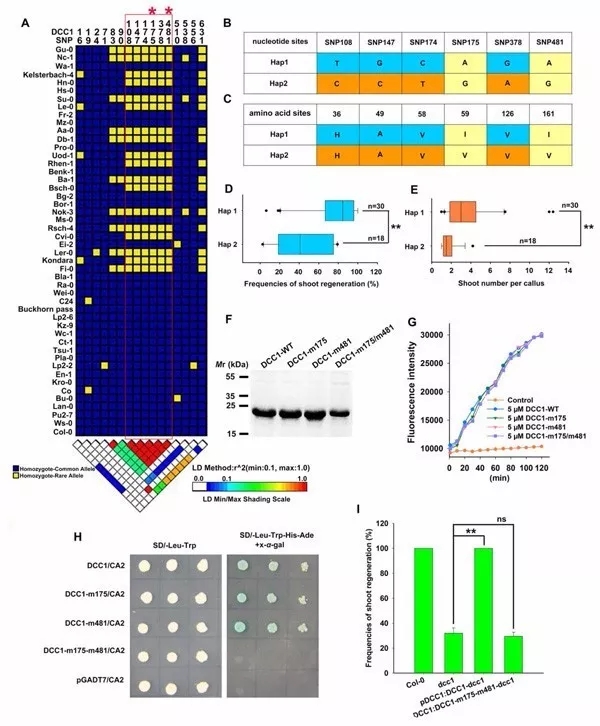
图8 DCC1调控不同生态型拟南芥芽再生能力
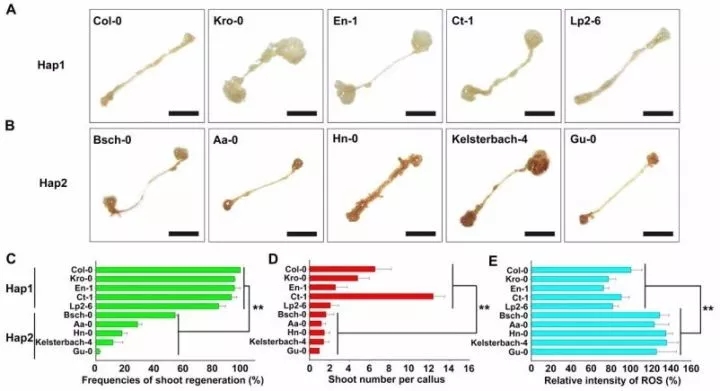
图9 不同生态型拟南芥的ROS水平与芽再生能力
总 结
DCC1氧化还原重塑CA2蛋白,调控Complex I活性,DCC1或CA2突变导致Complex I活性下调,产生ROS,ROS的上调抑制多个通路的基因表达,从而抑制芽再生。

该研究逻辑思路严谨,实验验证充分,揭示了硫氧还蛋白通过氧化还原重塑调控ROS内稳态,进而影响芽再生过程,这为理解植株再生的自然变异机制提供了新的证据。
参考文献
【1】Zhang H, Zhang T T, Liu H, et al. Thioredoxin-Mediated ROS Homeostasis Explains Natural Variation in Plant Regeneration.[J]. Plant Physiology, 2017:pp.00633.2017.
更多的原创文章,可继续关注我们网站动态发布,同时关注基迪奥微信~扫一扫添加基迪奥好友~随时随地关注行业动态!












